Are you one of those people who just can’t stand the idea of parasites? You find the idea of some slimy critter living in or on you and feeding on your very flesh just repulsive? If so, this post may be best viewed through a crack between fingers clamped tight across your eyes. If, on the other hand, you’re more like me and find the idea just fascinating (enough in my case to study them for grad school), then enjoy the following, which comprise a six-pack of the some of the best parasites you’ll find in fish. I’m not just restricting it to fish because this is a marine science blog, but because fish truly get some of the worst (best?) parasites around. Its probably got a lot to do with the diversity of available hosts (28,000+ species of fish = parasite smorgasboard!) and the supportive medium in which they live. Here they are then, six fish parasites you don’t want to miss.
1. Bite my tongue
Delightful little critters, tongue biters are a type of isopod, vaguely related to the pill bug/roly poly/slater bug you might see in a wood heap from time to time. They’re actually crustaceans more related to shrimp and crabs than to insects, so in a sense the common pill bug is more of a weirdo than the tongue biter; at least tongue biters have the decency to be truly aquatic, but I digress. Cymothoid isopods live on all sorts of fish in both fresh and salt water. They have a tick-like life-cycle where they periodically jump on and off the fish, taking a meal of blood or tissue each time they are on, and moulting each time they are off. Lots live on the skin or gills, and some even invade the body cavity, but the most famous are those that live in the mouth and hang onto the bottom lip, peering out over the teeth like a motorcyclist peeking over the windshield. They have quite good eyes, so they can probably see whats going on pretty well. Why live on the tongue? Not sure - oxygenated water supply perhaps? Maybe they just like the view. Whatever the reason, it would be a bit like you or I carrying a rhinoceros beetle around in your mouth all the time.
 Image credit: Australian Museum
Image credit: Australian Museum
2. Size does matter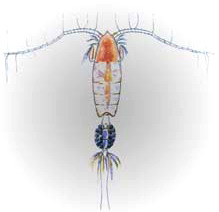 Image: NOAA
Image: NOAA
Your typical free-living planktonic copepod looks like the thing on the right here: tiny (<1mm or 1/20th in.), clearly crustacean, with delicate swimming antennae and one or two simple egg clumps hanging off the abdomen. Not so their parasitic cousins. In a pattern repeated often in the evolution of various parasite groups (parasitism having arisen in practically every phylum), the parasitic ones are much bigger than their free-living relatives. Why? It could be many reasons, like predator evasion, reproductive efficiency or just a result of living la dolce vita off some other poor sod. Whatever the reason, pennellid copepods take this size disparity to the extreme. In a group where the average size is less than 1mm, adult pennellids can be 30cm or more in length, even longer if you count the strands of eggs that trail behind them in the water like a string of pearls, popping little parasitic progeny off the end like pez. That’s three and half orders of magnitude in body size, and there aren’t many animal groups that can claim that sort of range. The largest pennellids look like big brown toilet b Image:Oceansunfish.orgrushes sticking out of the flesh of their hosts, which tend to be large pelagic things like sunfish, marlin and sharks (sometimes even dolphins!). The brush bit is the tail end and serves as a sort of gill, with countless tiny filaments to provide a high surface area and thereby facilitate oxygen exchange. If the brush is the back end, then that means the head is - you guessed it - buried deep in the flesh, usually T-ing into a blood vessel like a wall anchor and providing the copepod with a continuous source of food to fuel those ever-lengthening egg strands.
Image:Oceansunfish.orgrushes sticking out of the flesh of their hosts, which tend to be large pelagic things like sunfish, marlin and sharks (sometimes even dolphins!). The brush bit is the tail end and serves as a sort of gill, with countless tiny filaments to provide a high surface area and thereby facilitate oxygen exchange. If the brush is the back end, then that means the head is - you guessed it - buried deep in the flesh, usually T-ing into a blood vessel like a wall anchor and providing the copepod with a continuous source of food to fuel those ever-lengthening egg strands.
There’s a great scene in the BBC series Blue Planet (that paragon of marine biologist porn!) where a sea gull valiantly tries to remove pennellids from the skin of a basking sunfish, but in vain. It proves nicely that a large body size confers on a parasite both robustness and protection from predators. Its one of the best and most remarkable scenes in the entire film series.
3. Going to great lengths
 This isn’t Nematobibothrioides, but a similar species from Hawaiian jobfish. Image:HIMBThe ocean sunfish, Mola mola, is also host to the third in our set: the didymozoid Nematobibothrioides histoidii. I know, awful names, right? Didymozoids (pronounced like P-Diddy) are a family of parasitic flatworms in the digenean (“fluke”) group. Most digeneans live in the gut, but didymozoids have invaded the flesh. That’s not what makes this worm special, though, there are lots of worms that live in flesh. No, its the length of N. histoidii that sets it apart. In the best paper on the subject Glenn Noble (1975) describes finding them up to 12 meters long! That’s a 40 foot worm, as Noble says “snaking” its way through the tissues, leaving a trail of eggs in its wake. Chances are, N. histoidii may even grow larger than that. Scientists don’t know, partly because dissections of Mola aren’t all that common and partly because dissecting an entire sunfish to extract the fragile, threadlike worm in one piece is practically impossible. Didymozoids have incredibly complex life cycles that likely involve flaoting snails and copepods and all sorts of other intermediate hosts. Despite this, or perhaps because of it, they’ve been very successful in infecting a wide range of pelagic fishes. I probably shouldn’t tell you that a significant proportion of the tuna you’ve ever eaten probably hosted didymozoids somewhere in its body when it was alive. Oh wait, I just did…
This isn’t Nematobibothrioides, but a similar species from Hawaiian jobfish. Image:HIMBThe ocean sunfish, Mola mola, is also host to the third in our set: the didymozoid Nematobibothrioides histoidii. I know, awful names, right? Didymozoids (pronounced like P-Diddy) are a family of parasitic flatworms in the digenean (“fluke”) group. Most digeneans live in the gut, but didymozoids have invaded the flesh. That’s not what makes this worm special, though, there are lots of worms that live in flesh. No, its the length of N. histoidii that sets it apart. In the best paper on the subject Glenn Noble (1975) describes finding them up to 12 meters long! That’s a 40 foot worm, as Noble says “snaking” its way through the tissues, leaving a trail of eggs in its wake. Chances are, N. histoidii may even grow larger than that. Scientists don’t know, partly because dissections of Mola aren’t all that common and partly because dissecting an entire sunfish to extract the fragile, threadlike worm in one piece is practically impossible. Didymozoids have incredibly complex life cycles that likely involve flaoting snails and copepods and all sorts of other intermediate hosts. Despite this, or perhaps because of it, they’ve been very successful in infecting a wide range of pelagic fishes. I probably shouldn’t tell you that a significant proportion of the tuna you’ve ever eaten probably hosted didymozoids somewhere in its body when it was alive. Oh wait, I just did…
4. Beauty in the beast
Trichodina is my favourite protistan (single-celled) parasite; it’s one that parasitology students and researchers alike are drawn to for its spectacular marriage of form and function and for the startling and beautiful complexity inherent in a single cell. Trichodinids are ciliated parasites of the skin, fins and gills (mostly) of fishes (mostly) in both fresh and salt water, and they’re shaped as discs or hemispheres, sucking onto the surface of the fish with their flattened underside. Everyone always shouts “Scubbing bubbles!” when I show them in parasitology classes, and I guess I can see why:
 Trichodina and Scrubbing Bubbles - separated at birth?
Trichodina and Scrubbing Bubbles - separated at birth?
What makes Trichodina so amazing is a ring of interlocking structures just under the cell membrane, called denticles, that are used to aid in attachment to wet fish skin, which is pretty slippery stuff. When the pins in the middle of the denticle pull up, the blades on the outside bite down into the skin, with the added bonus of creating suction on the underside. The blades are revealed by staining the cells in silver nitrate and then exposing them to UV light to develop them, exactly like a photograph, and their structure is very important for telling species apart, which is good, because there’s over 150 in the genus! Some of them are definitely parasitic and can cause nasty disease, but most are  probably commensals, which means they’re just using the host as a home and means of conveyance while they feed on bacteria and other detritus suspended in the water. One of the most captivating things I ever saw was a trichodinid cell dividing, under a microscope. The way a single-cell could disengage all those blades (each daughter cell gets half), successfully divide and then replicate the missing denticles and reassemble their intricate structure was just mesmerizing.
probably commensals, which means they’re just using the host as a home and means of conveyance while they feed on bacteria and other detritus suspended in the water. One of the most captivating things I ever saw was a trichodinid cell dividing, under a microscope. The way a single-cell could disengage all those blades (each daughter cell gets half), successfully divide and then replicate the missing denticles and reassemble their intricate structure was just mesmerizing.
5. Stop it, or you’ll go blind
 Diplostome metacercariae in a fish’s lens. http://www.thefishsite.com/ Eyes are sensitive things, so the thought of some freeloading bug making a home in your precious orbits is just creepy, and yet parasites in the eyes are pretty common. Why? There’s several possibilities, but the two best are, firstly, that the eye is a relatively inactive place as far as the immune response goes (not a lot of blood inside your eyeball, normally anyway) and that, secondly, infecting the eyes can help you get where you’re going next. If we, ahem, focus on the second reason for a minute, I can tell you about diplostomes. These are another family of digenean flatworms, but unlike Nematobibothrioides in the Mola, these mature in birds - fish-eating birds to be exact. The stage that infects fish is an immature form that lives in the lenses of the eye. They’re very common, almost ubiquitous, among freshwater fishes, but nearly always only in one eye. If we, um, look at some fish and find that a third of individuals have diplostomes in the left eye and third of fish have them in the right eye, then a ninth of fish (1/3 x 1/3 = 1/9) should have it in both eyes, but this is not what we, ahem, see. Where are the dual infections? You guessed it - nailed, um, in the blink of an eye by one of the aforementioned piscivorous birds. A fish, it seems, can get by with being functionally blinded in one eye, but being blind in both makes you, er, a sight for sore eyes to your average heron. In this way the parasite is playing probability statistics to get its life-cycle completed; worms are good at math, who knew?
Diplostome metacercariae in a fish’s lens. http://www.thefishsite.com/ Eyes are sensitive things, so the thought of some freeloading bug making a home in your precious orbits is just creepy, and yet parasites in the eyes are pretty common. Why? There’s several possibilities, but the two best are, firstly, that the eye is a relatively inactive place as far as the immune response goes (not a lot of blood inside your eyeball, normally anyway) and that, secondly, infecting the eyes can help you get where you’re going next. If we, ahem, focus on the second reason for a minute, I can tell you about diplostomes. These are another family of digenean flatworms, but unlike Nematobibothrioides in the Mola, these mature in birds - fish-eating birds to be exact. The stage that infects fish is an immature form that lives in the lenses of the eye. They’re very common, almost ubiquitous, among freshwater fishes, but nearly always only in one eye. If we, um, look at some fish and find that a third of individuals have diplostomes in the left eye and third of fish have them in the right eye, then a ninth of fish (1/3 x 1/3 = 1/9) should have it in both eyes, but this is not what we, ahem, see. Where are the dual infections? You guessed it - nailed, um, in the blink of an eye by one of the aforementioned piscivorous birds. A fish, it seems, can get by with being functionally blinded in one eye, but being blind in both makes you, er, a sight for sore eyes to your average heron. In this way the parasite is playing probability statistics to get its life-cycle completed; worms are good at math, who knew?
6. When the worm turns
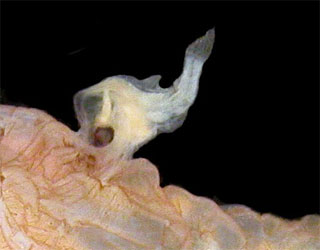
Our first 5 candidates were all parasites OF fish, but the title of the post was “Six fish parasites” and that could just as easily mean the fish IS the parasite. I thought about doing the male anglerfish, which parasitises his mate so intimately that their circulatory systems fuse, or the candiru, that tiny terror of the Amazon: a catfish that swims into the human urethra and locks its spines erect, making removal a matter for a skilled surgeon. But in the, um, end, I had to go with the pearlfish (I’m sorry, I’ll stop now, promise). Thats because this delightful fish, rather humourously called Carapus, chooses to live in the lower digestive system of sea cucumbers, a lumbering sausage-like relative of urchins and starfish that spends its days lazily sifting food particles from sand on coral reefs. Pearlfish may not, strictly speaking, be parasites, since it seems that they feed while on day-trip excursions out of the sphincter, rather than on the cucumber itself, but I’m told they can and do feed on the respiratory tree of the host if the need arises. Its a good thing sea cucumbers are masters of regeneration. I’ve never actually seen one, but then again I dont spend a lot of time peering into beche-de-mer butts
 Image: Claude Rives/Fishbase
Image: Claude Rives/Fishbase
Its often said that parasitism is the most common lifestyle on the planet, and nowhere is this better seen than among fish hosts. From forty foot flukes to tongue-biting isopods, fish are home to the most amazing variety of parasitic symbionts. Estimates of how many species of fish parasites there are run into the hundreds of thousands; vastly more than the diversity of fish, which are themselves the most diverse vertebrate phylum. Next time you look at a fish, then, try to see it for what it really is: a little swimming city, replete with enclaves of surprising parasite diversity in practically every tissue.
 Camouflaged brittle star attached to a gorgonian
Camouflaged brittle star attached to a gorgonian
 Share Article
Share Article 












{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}